

Disclaimer:
The authors are solely responsible for the content of this report. Material included herein does not represent the opinion of the European Community, and the European Community is not responsible for any use that might be made of it.
Back to overview reports
The importance of these estuarine areas for bird populations is mainly due to the availability of a mosaic of habitats (intertidal mudflats and sandflats, marshes, grasslands) for feeding and/or roosting, leading to the designation of substantial parts of these areas as Special Protection Areas, under the European Birds Directive, Special Conservation Areas, under the Habitats and Species Directive, as well as Ramsar sites because of their international importance as wetlands. However, these estuaries are also places of intense human activities (e.g. port activities, water abstraction, fishery, habitat claim) thus leading to the presence of several conflicts with the conservation of these sites as bird habitats (see TIDE report “Analysis of the TIDE estuarine conflict matrices”). A key element for the management of these conflicts (e.g. through provision of appropriate mitigation and compensation of the impacts arising from human activities) is the knowledge of the distribution of bird species within these sites and, in particular, the understanding of the critical determinants affecting their use of estuarine habitats. With this purpose, the influence of several environmental characteristics (covering aspects such as habitat availability and type, quality of the feeding area, level of anthropogenic disturbance) on the distribution of bird species within the Humber, Weser and Elbe estuaries has been investigated.
The studied estuaries are characterised by broadly similar conditions (e.g. strong tidal influence, transport of large quantities of sediment, presence of large port areas), and a broadly similar distribution of bird species within these areas has been observed, with higher bird abundance generally present in the outer parts of these estuaries, where large intertidal feeding areas are often present (this is particularly true in the Humber and Weser estuaries) and species using also other coastal habitats occur with higher frequency and abundance. Higher waterbird abundances in the polyhaline and mesohaline zones have also been reported in the Scheldt estuary (Ysebaert et al. 2000). However, a certain variability in the species-habitat association has been observed among the studied estuaries (e.g. with the oligohaline zone in the Humber showing a higher relevance, particularly to waders, compared to similar salinity zones in the other estuaries), highlighting the importance of local conditions in affecting habitat use by birds. The habitat tolerances of most wader species, in fact, may be fairly broad, and a certain degree of opportunism is present (Prater 1981, McLusky and Elliott 2004), thus leading to adaptations to local conditions due e.g. to the different distribution and availability of food resources in the estuary. In addition, as shown by the multivariate and univariate models applied in this study, it is usually the combination of several variables that affects the species distribution in an estuarine area rather than a single factor alone (although certain variables may have a higher relevance than others), thus highlighting the complexity of the species-habitat relationship.
In general, an overall positive relationship has been observed between bird species densities and the habitat area, in particular the intertidal area, suggesting that larger mudflats might have a greater carrying capacity per unit of area. The size of any productive area in an estuary is generally positively associated to its carrying capacity in supporting wading birds, in terms of maximum number of individuals (or biomass) that can be sustained (Meire 1993, Elliott et al. 1998). However, when the density of individuals in the estuarine area is considered (i.e., the number of individuals per unit area), a lower wader density has been reported in larger estuarine areas, this negative relationship possibly ascribed to the inclusion of many unsuitable feeding areas (e.g. deeper subtidal areas) in these cases (Prater 1981). Although this explanation may be valid at the larger inter-estuarine scale, a different one might support the opposite pattern at the smaller intra-estuarine scale as observed in the present study, particularly when considering the area of suitable feeding habitats such as intertidal mudflats. Given that food is considered to be the major determinant of shorebird distribution (Prater 1981, McLusky and Elliott 2004), the relationship with the intertidal habitat area may be linked to the availability of food resources in it. In particular, wider, more extensive habitat areas are likely to have a higher diversity of microhabitats (hence a possible higher diversity in the food resources) and this might lead to a higher probability for bird species of accessing different food resources, possibly resulting also in a reduction in the possible intra- and inter-specific competition, thus allowing a higher concentration of individuals in larger habitat areas. However, it is acknowledged that habitat size alone will not necessarily determine wader distribution, with other site specific factors also influencing this.
This relationship is likely to be particularly relevant to generalist mudflat feeders, as observed in the case of Dunlin, Redshank and Shelduck. However, these species showed a different preference for the type of intertidal habitat, with Shelduck density being associated mostly to substrata dominated by littoral mud, whereas Redshank occurring in greater densities where a littoral sand component is also present in the substratum. Dunlin and Redshank feed throughout the estuary on marine polychaete worms, crustaceans and molluscs, such as the Baltic Tellin Macoma balthica, tending to be near the water’s edge (McLusky and Elliott 2004). A positive effect of intertidal habitat area on these species density has been observed in the Humber and Weser (for Dunlin) (particularly with intertidal area >3 km2), with higher concentrations of the species in the outer estuarine areas, where larger counting units (hence larger habitat areas) are present. In addition the presence of wider suitable habitats is likely to provide unrestricted views (compared, for example, to narrower mudflats in the upper Humber estuary) for the early detection of predators. It is of note that although Redshank can be considered a generalist feeder, it also shows some preference for Corophium in many estuaries (including the Humber), this invertebrate occurring mainly on the upper and mid shore flats, where mostly Redshank feed (Prater 1981). Higher density of Redshank is also related in the Humber to muddy sand / sandy mud substrata with general high total benthic abundance.
It is of note that an opposite (negative) relationship of Dunlin density with the habitat area has been observed in the Elbe estuary, this result likely being an artefact of the analysis being influenced by the data obtained from the polyhaline zone of this estuary (e7NDS), regarding outer sands and remote islands in the Wadden Sea. In fact, in this zone there is the combination of very small counting units (max. 0.2 km2, hence small area of the habitats therein) compared to those in the other zones of the estuary (with a minimum area between 2 and 8 km2) and extremely high counts, hence very high density, of Dunlin, particularly around the island of Scharhöm compared to the other areas, thus driving the negative relationship of the species density with habitat areas for the whole estuary. It is of note also that when the effect of habitat area is excluded, the relationship with the salinity gradient is still relevant to Dunlin density in the Elbe estuary (with salinity zone included in the final model) with higher Dunlin density expected in oligohaline and mesohaline zones. This suggests that factors other than salinity and habitat availability within the single units are the likely determinants of the very high density values observed for Dunlin in the polyhaline zone of the Elbe, and it might be expected that there will be a positive relationship between the availability of foraging area and roost size in many instances, flocks tending to minimize flight distance between preferred foraging and roosting areas where possible.
In turn, the higher diversity of microhabitat (and the associated food resources) is likely not to have a positive effect on specialist feeders, the relationship between the habitat area and the bird density being dependent on the availability of specific prey/microhabitat. An example is Bar-tailed Godwit in the Humber, showing higher density in sectors with smaller intertidal area (<5 km2). Although, like Dunlin and Redshank, this species feeds on benthic prey available on estuarine mudflats, it shows a higher degree of specialism, feeding on larger prey (e.g. large polychaetes and bivalves; Scheiffarth 2001). Due to their long bill, Bar-tailed Godwit can access to larger prey that usually bury themselves more deeply than do smaller individuals (as in the case of larger Macoma balthica) (Prater 1981). Different studies have highlighted the presence of larger Macoma individuals in the lower intertidal compared to the upper intertidal zone (Bouma et al. 2001, Hiddink et al. 2002), the former habitat likely being more readily accessible from the shore roosting sites where the mudflat is smaller (and possibly narrower) compared to wider mudflats. The negative relationship observed between Bar-tailed Godwit density and the total benthic abundance in the intertidal habitat is also likely to support the preference of the species for feeding habitats where benthic communities are dominated by larger prey (usually in lower densities compared to smaller invertebrates).
Higher density of Golden Plover is also predicted in the Humber where larger intertidal marsh habitats (>0.6 km2) occur, in combination with smaller subtidal areas (<10 km2). The association of higher densities of the species with a particular intertidal habitat type (littoral sand) and benthic community was also observed, but this cannot be explained by the feeding preferences of the species. Golden Plover, in fact, use estuarine habitat mainly for roosting, with feeding primarily on inland habitats (e.g. habitats with short vegetation, like bare peats, wet vegetation, pasture fields with short swards).
When investigating the occurrence of two wildfowl species, namely Pochard and Brent Goose in the Humber Estuary, different relationships with habitat areas are observed, as a result of the different feeding habitats of these two species. Pochard is a freshwater duck, with omnivorous feeding habits, the analysis identifying its most suitable habitat within the estuary to be where there is a higher ratio between marsh and intertidal area (i.e. marsh area >0.84 km2 and intertidal area <10 km2), the former habitat likely to be preferred by the species due to the diversity and abundance of feeding resources (including seeds, roots, rhizomes and the vegetative parts of grasses, sedges and aquatic plants, as well as aquatic insects and larvae, molluscs, crustaceans, worms, amphibians and small fish). In turn, the Brent Goose feeds almost exclusively on the Zostera/Enteromorpha beds, this food resource being likely more available where larger intertidal areas (>10 km2) occur.
The distribution of anthropogenic activities along the estuarine banks proved to have a relevant effect on the distribution of bird species, generally resulting in a lower density of birds in areas where the estuarine bank has been modified through the creation of artificial hard substrata (e.g. docks and seawalls in sectors ND and NE in the Humber) or where industrial sites and other infrastructures are concentrated (like in the Elbe estuary, leading to differences between the north and south bank within similar salinity zones of the estuary). A specific index of anthropogenic disturbance has been included in the analysis for the Humber estuary. However, although a negative effect of anthropogenic disturbance would be expected on bird densities, a positive relationship between the bird density and the index used was often observed (e.g. for Dunlin, Redshank and Shelduck). Although Dunlin, Redshank and Shelduck may be fairly tolerant to disturbance from recreation (English Nature 2003), it is likely that the above result is an artefact of the analysis deriving from the inadequacy of the measured index as a proxy for disturbance given the spatial scale considered by the analysis.
In fact, although the index accounts for the frequency of potentially disturbing activities in the estuarine areas (including shore-based, water-based and airborne activities), this might not correspond to an actual disturbance to bird populations roosting within a sector. Indeed, it might be expected that roost sites are actively identified by birds in areas where disturbance is at a low level. Therefore, there could be a mismatch between the location of the disturbing activity and the location of roosting/compression sites within a sector, this mismatch not being captured by the analysis, as the sector as a whole was used as the minimum spatial unit. In addition, there might be other correlated factors, i.e. showing a similar distribution among sectors as disturbance, but not measured here, that might have contributed to this result. All these elements may have led to artefacts in the analysis resulting in the apparent positive effect of disturbance level on Dunlin, Redshank and Shelduck density.
The data availability for the water quality in the studied estuaries limits the interpretation of the obtained results to the Elbe estuary and to the freshwater and oligohaline zone of the Weser. Water quality characteristics (including indicators of the salinity gradient, nutrient levels, organic enrichment) resulted more important in affecting bird assemblages as a whole in the Elbe compared to the Weser, although this comparison must be taken with caution, due to the different datasets analysed and also the differences in their spatial coverage of the estuarine area. The effect of the salinity gradient was predominant in the Elbe (where data covered the whole salinity gradient) when considering the density of bird assemblages as a whole, as well as the occurrence of Dunlin. In this site, as in the rest of the Wadden Sea, other species show a strong association with salinity, as with the Brent Goose, usually concentrating on the islands and the outer coast. The principal effect of the changes in the salinity regime is usually related to changes in communities of benthic invertebrates (Cole et al. 1999). However, the results for the Elbe might be influenced by an artefact of the analysis, due to the small size of counting units in the outer (polyhaline) estuary where high bird counts were recorded, leading to very high bird densities as described before. PO4 also proved to be a relevant predictor of wader and wildfowl overall distribution in the Elbe, with higher densities of most of wader species (including Dunlin and Redshank) and of several wildfowl species (including Shelduck and Brent Goose) recorded in the outer estuary where PO4 levels are lower. The probability of occurrence of Dunlin in the estuary was also higher at lower PO4 concentration (<0.002 mmol/l). Nutrient and organic enrichment can lead to an increase in benthic populations such as opportunistic marine worms, whereas a decline in the input of nutrients may correlate with lower phytoplankton biomass, resulting in a decline of the stocks of filter feeding bivalves and a consequent decline in shellfish eaters, as observed in the western Dutch Wadden Sea (van Roomen et al. 2012). However, as regards the results obtained for the Elbe estuary, there might be a relevant effect of the very high bird densities resulting in the polyhaline zone of the Elbe estuary, as explained before, where the lowest PO4 levels are lower, thus suggesting the influence of other factors not included in the analysis.
Finally, it should be noted that the interpretation of the results described above in terms of bird habitat use might be limited by the fact that high-tide counts were used to derive the analysed species densities in the estuary. These counts, are carried out when most of the preferred intertidal feeding habitats are temporarily unavailable to the species, with most either foraging on the upper shore in sub-optimal areas or roosting in the area while waiting for the tide to retreat. A certain site fidelity of the species was assumed, hence considering the obtained density data as representative of the bird use of the area, as most species tend to simply move up and/or along shore during tidal compression (and depending on tide), with roosts usually located as close to preferred foraging areas as possible. However, it must be acknowledged that the use of high-tide counts might lead to an underestimation of the birds using an area at low tide, particularly for those species (e.g. Redshank, Curlew and Oystercatcher) which can move inland in search of food when their estuarine food resources are not accessible (Prater 1981). Using low-tide counts is likely to allow better relationships of the actual density with the habitat availability and characteristics in the counting unit areas, although this type of data might present other limitations compared to the high-tide counts. For example, in the Humber, low-tide counts are carried out at a lower frequency (e.g. c. every 5 years) hence limiting the availability of data for the statistical analysis.
Symbols + and – indicate a general positive or negative relationship with the relevant environmental descriptor (thresholds and ranges identifying optimal conditions for the species occurrence/density are provided in brackets). +/- indicates a fluctuating relationship (when no clear increase or decrease pattern can be identified; the shape of these relationships being shown in Chapter 6). When symbols are within brackets, these indicate predictors that have not been included in the models because of their correlation with other variables included, but, due to their collinearity with the relevant descriptors, their influence cannot be excluded.
Back to top
What environmental factors should be considered in the design of a compensation scheme for waterbirds and their habitats?
What environmental variables are most important in determining minimizing or basic compensatory requirements for waterbirds?
What is important in establishing a zonation for estuaries?
What tools and guidance are available to minimise and mitigate disturbance to waterbirds?
Determinants of bird habitat use in TIDE estuaries
Table of content
- 1. SUMMARY
- 2. INTRODUCTION
- 3. STRUCTURE OF THE REPORT
- 4. DATA USED
- 5. GENERAL CHARACTERISTICS OF BIRD ASSEMBLAGES IN TIDE ESTUARIES
- 6. BIRD ASSEMBLAGES DISTRIBUTION AND RELATIONSHIP WITH ENVIRONMENTAL VARIABLES
- 6a. Humber
- 6b. Weser
- 6c. Elbe
- 7. SPECIES DISTRIBUTION MODELS
- 7a. Dunlin
- 7b. Redshank, Golden Plover and Bar-tailed Godwit
- 7c. Shelduck, Pochard and Brent Goose
- 8. DISCUSSION
- 9. CONCLUSIONS
- 9a. Analysis Conclusions
- 9b. Management Recommendations
- 9c. Recommendations for Future Studies
- 10. REFERENCES
- 11. APPENDIX 1
- 12. APPENDIX 2
- 13. APPENDIX 3
- 14. APPENDIX 4
8. Discussion
The Humber, Weser and Elbe estuaries are important sites for the conservation of bird populations. They are essential components in a network of wetland sites constituting the East Atlantic Flyway stretching from the Arctic Circle, to southern Europe, west Africa and sometimes as far as southern Africa, providing migratory birds with suitable feeding and resting habitats during the great migrations between northern breeding grounds and southern wintering sites (English Nature 2003, McLusky and Elliott 2004, van Roomen et al. 2012). As a result, these estuaries support internationally important populations (i.e. where there is a regular occurrence of at least 1% of their flyway or biogeographical population) of several species, for example Dark-bellied Brent Goose, Shelduck, Golden Plover, Lapwing and Knot in the case of the Humber (English Nature 2003), making this estuary one of the top five most important wetland sites in the UK, and the top ten in Europe, for the population of over-wintering and migratory birds which depend on it.The importance of these estuarine areas for bird populations is mainly due to the availability of a mosaic of habitats (intertidal mudflats and sandflats, marshes, grasslands) for feeding and/or roosting, leading to the designation of substantial parts of these areas as Special Protection Areas, under the European Birds Directive, Special Conservation Areas, under the Habitats and Species Directive, as well as Ramsar sites because of their international importance as wetlands. However, these estuaries are also places of intense human activities (e.g. port activities, water abstraction, fishery, habitat claim) thus leading to the presence of several conflicts with the conservation of these sites as bird habitats (see TIDE report “Analysis of the TIDE estuarine conflict matrices”). A key element for the management of these conflicts (e.g. through provision of appropriate mitigation and compensation of the impacts arising from human activities) is the knowledge of the distribution of bird species within these sites and, in particular, the understanding of the critical determinants affecting their use of estuarine habitats. With this purpose, the influence of several environmental characteristics (covering aspects such as habitat availability and type, quality of the feeding area, level of anthropogenic disturbance) on the distribution of bird species within the Humber, Weser and Elbe estuaries has been investigated.
The studied estuaries are characterised by broadly similar conditions (e.g. strong tidal influence, transport of large quantities of sediment, presence of large port areas), and a broadly similar distribution of bird species within these areas has been observed, with higher bird abundance generally present in the outer parts of these estuaries, where large intertidal feeding areas are often present (this is particularly true in the Humber and Weser estuaries) and species using also other coastal habitats occur with higher frequency and abundance. Higher waterbird abundances in the polyhaline and mesohaline zones have also been reported in the Scheldt estuary (Ysebaert et al. 2000). However, a certain variability in the species-habitat association has been observed among the studied estuaries (e.g. with the oligohaline zone in the Humber showing a higher relevance, particularly to waders, compared to similar salinity zones in the other estuaries), highlighting the importance of local conditions in affecting habitat use by birds. The habitat tolerances of most wader species, in fact, may be fairly broad, and a certain degree of opportunism is present (Prater 1981, McLusky and Elliott 2004), thus leading to adaptations to local conditions due e.g. to the different distribution and availability of food resources in the estuary. In addition, as shown by the multivariate and univariate models applied in this study, it is usually the combination of several variables that affects the species distribution in an estuarine area rather than a single factor alone (although certain variables may have a higher relevance than others), thus highlighting the complexity of the species-habitat relationship.
In general, an overall positive relationship has been observed between bird species densities and the habitat area, in particular the intertidal area, suggesting that larger mudflats might have a greater carrying capacity per unit of area. The size of any productive area in an estuary is generally positively associated to its carrying capacity in supporting wading birds, in terms of maximum number of individuals (or biomass) that can be sustained (Meire 1993, Elliott et al. 1998). However, when the density of individuals in the estuarine area is considered (i.e., the number of individuals per unit area), a lower wader density has been reported in larger estuarine areas, this negative relationship possibly ascribed to the inclusion of many unsuitable feeding areas (e.g. deeper subtidal areas) in these cases (Prater 1981). Although this explanation may be valid at the larger inter-estuarine scale, a different one might support the opposite pattern at the smaller intra-estuarine scale as observed in the present study, particularly when considering the area of suitable feeding habitats such as intertidal mudflats. Given that food is considered to be the major determinant of shorebird distribution (Prater 1981, McLusky and Elliott 2004), the relationship with the intertidal habitat area may be linked to the availability of food resources in it. In particular, wider, more extensive habitat areas are likely to have a higher diversity of microhabitats (hence a possible higher diversity in the food resources) and this might lead to a higher probability for bird species of accessing different food resources, possibly resulting also in a reduction in the possible intra- and inter-specific competition, thus allowing a higher concentration of individuals in larger habitat areas. However, it is acknowledged that habitat size alone will not necessarily determine wader distribution, with other site specific factors also influencing this.
This relationship is likely to be particularly relevant to generalist mudflat feeders, as observed in the case of Dunlin, Redshank and Shelduck. However, these species showed a different preference for the type of intertidal habitat, with Shelduck density being associated mostly to substrata dominated by littoral mud, whereas Redshank occurring in greater densities where a littoral sand component is also present in the substratum. Dunlin and Redshank feed throughout the estuary on marine polychaete worms, crustaceans and molluscs, such as the Baltic Tellin Macoma balthica, tending to be near the water’s edge (McLusky and Elliott 2004). A positive effect of intertidal habitat area on these species density has been observed in the Humber and Weser (for Dunlin) (particularly with intertidal area >3 km2), with higher concentrations of the species in the outer estuarine areas, where larger counting units (hence larger habitat areas) are present. In addition the presence of wider suitable habitats is likely to provide unrestricted views (compared, for example, to narrower mudflats in the upper Humber estuary) for the early detection of predators. It is of note that although Redshank can be considered a generalist feeder, it also shows some preference for Corophium in many estuaries (including the Humber), this invertebrate occurring mainly on the upper and mid shore flats, where mostly Redshank feed (Prater 1981). Higher density of Redshank is also related in the Humber to muddy sand / sandy mud substrata with general high total benthic abundance.
It is of note that an opposite (negative) relationship of Dunlin density with the habitat area has been observed in the Elbe estuary, this result likely being an artefact of the analysis being influenced by the data obtained from the polyhaline zone of this estuary (e7NDS), regarding outer sands and remote islands in the Wadden Sea. In fact, in this zone there is the combination of very small counting units (max. 0.2 km2, hence small area of the habitats therein) compared to those in the other zones of the estuary (with a minimum area between 2 and 8 km2) and extremely high counts, hence very high density, of Dunlin, particularly around the island of Scharhöm compared to the other areas, thus driving the negative relationship of the species density with habitat areas for the whole estuary. It is of note also that when the effect of habitat area is excluded, the relationship with the salinity gradient is still relevant to Dunlin density in the Elbe estuary (with salinity zone included in the final model) with higher Dunlin density expected in oligohaline and mesohaline zones. This suggests that factors other than salinity and habitat availability within the single units are the likely determinants of the very high density values observed for Dunlin in the polyhaline zone of the Elbe, and it might be expected that there will be a positive relationship between the availability of foraging area and roost size in many instances, flocks tending to minimize flight distance between preferred foraging and roosting areas where possible.
In turn, the higher diversity of microhabitat (and the associated food resources) is likely not to have a positive effect on specialist feeders, the relationship between the habitat area and the bird density being dependent on the availability of specific prey/microhabitat. An example is Bar-tailed Godwit in the Humber, showing higher density in sectors with smaller intertidal area (<5 km2). Although, like Dunlin and Redshank, this species feeds on benthic prey available on estuarine mudflats, it shows a higher degree of specialism, feeding on larger prey (e.g. large polychaetes and bivalves; Scheiffarth 2001). Due to their long bill, Bar-tailed Godwit can access to larger prey that usually bury themselves more deeply than do smaller individuals (as in the case of larger Macoma balthica) (Prater 1981). Different studies have highlighted the presence of larger Macoma individuals in the lower intertidal compared to the upper intertidal zone (Bouma et al. 2001, Hiddink et al. 2002), the former habitat likely being more readily accessible from the shore roosting sites where the mudflat is smaller (and possibly narrower) compared to wider mudflats. The negative relationship observed between Bar-tailed Godwit density and the total benthic abundance in the intertidal habitat is also likely to support the preference of the species for feeding habitats where benthic communities are dominated by larger prey (usually in lower densities compared to smaller invertebrates).
Higher density of Golden Plover is also predicted in the Humber where larger intertidal marsh habitats (>0.6 km2) occur, in combination with smaller subtidal areas (<10 km2). The association of higher densities of the species with a particular intertidal habitat type (littoral sand) and benthic community was also observed, but this cannot be explained by the feeding preferences of the species. Golden Plover, in fact, use estuarine habitat mainly for roosting, with feeding primarily on inland habitats (e.g. habitats with short vegetation, like bare peats, wet vegetation, pasture fields with short swards).
When investigating the occurrence of two wildfowl species, namely Pochard and Brent Goose in the Humber Estuary, different relationships with habitat areas are observed, as a result of the different feeding habitats of these two species. Pochard is a freshwater duck, with omnivorous feeding habits, the analysis identifying its most suitable habitat within the estuary to be where there is a higher ratio between marsh and intertidal area (i.e. marsh area >0.84 km2 and intertidal area <10 km2), the former habitat likely to be preferred by the species due to the diversity and abundance of feeding resources (including seeds, roots, rhizomes and the vegetative parts of grasses, sedges and aquatic plants, as well as aquatic insects and larvae, molluscs, crustaceans, worms, amphibians and small fish). In turn, the Brent Goose feeds almost exclusively on the Zostera/Enteromorpha beds, this food resource being likely more available where larger intertidal areas (>10 km2) occur.
The distribution of anthropogenic activities along the estuarine banks proved to have a relevant effect on the distribution of bird species, generally resulting in a lower density of birds in areas where the estuarine bank has been modified through the creation of artificial hard substrata (e.g. docks and seawalls in sectors ND and NE in the Humber) or where industrial sites and other infrastructures are concentrated (like in the Elbe estuary, leading to differences between the north and south bank within similar salinity zones of the estuary). A specific index of anthropogenic disturbance has been included in the analysis for the Humber estuary. However, although a negative effect of anthropogenic disturbance would be expected on bird densities, a positive relationship between the bird density and the index used was often observed (e.g. for Dunlin, Redshank and Shelduck). Although Dunlin, Redshank and Shelduck may be fairly tolerant to disturbance from recreation (English Nature 2003), it is likely that the above result is an artefact of the analysis deriving from the inadequacy of the measured index as a proxy for disturbance given the spatial scale considered by the analysis.
In fact, although the index accounts for the frequency of potentially disturbing activities in the estuarine areas (including shore-based, water-based and airborne activities), this might not correspond to an actual disturbance to bird populations roosting within a sector. Indeed, it might be expected that roost sites are actively identified by birds in areas where disturbance is at a low level. Therefore, there could be a mismatch between the location of the disturbing activity and the location of roosting/compression sites within a sector, this mismatch not being captured by the analysis, as the sector as a whole was used as the minimum spatial unit. In addition, there might be other correlated factors, i.e. showing a similar distribution among sectors as disturbance, but not measured here, that might have contributed to this result. All these elements may have led to artefacts in the analysis resulting in the apparent positive effect of disturbance level on Dunlin, Redshank and Shelduck density.
The data availability for the water quality in the studied estuaries limits the interpretation of the obtained results to the Elbe estuary and to the freshwater and oligohaline zone of the Weser. Water quality characteristics (including indicators of the salinity gradient, nutrient levels, organic enrichment) resulted more important in affecting bird assemblages as a whole in the Elbe compared to the Weser, although this comparison must be taken with caution, due to the different datasets analysed and also the differences in their spatial coverage of the estuarine area. The effect of the salinity gradient was predominant in the Elbe (where data covered the whole salinity gradient) when considering the density of bird assemblages as a whole, as well as the occurrence of Dunlin. In this site, as in the rest of the Wadden Sea, other species show a strong association with salinity, as with the Brent Goose, usually concentrating on the islands and the outer coast. The principal effect of the changes in the salinity regime is usually related to changes in communities of benthic invertebrates (Cole et al. 1999). However, the results for the Elbe might be influenced by an artefact of the analysis, due to the small size of counting units in the outer (polyhaline) estuary where high bird counts were recorded, leading to very high bird densities as described before. PO4 also proved to be a relevant predictor of wader and wildfowl overall distribution in the Elbe, with higher densities of most of wader species (including Dunlin and Redshank) and of several wildfowl species (including Shelduck and Brent Goose) recorded in the outer estuary where PO4 levels are lower. The probability of occurrence of Dunlin in the estuary was also higher at lower PO4 concentration (<0.002 mmol/l). Nutrient and organic enrichment can lead to an increase in benthic populations such as opportunistic marine worms, whereas a decline in the input of nutrients may correlate with lower phytoplankton biomass, resulting in a decline of the stocks of filter feeding bivalves and a consequent decline in shellfish eaters, as observed in the western Dutch Wadden Sea (van Roomen et al. 2012). However, as regards the results obtained for the Elbe estuary, there might be a relevant effect of the very high bird densities resulting in the polyhaline zone of the Elbe estuary, as explained before, where the lowest PO4 levels are lower, thus suggesting the influence of other factors not included in the analysis.
Finally, it should be noted that the interpretation of the results described above in terms of bird habitat use might be limited by the fact that high-tide counts were used to derive the analysed species densities in the estuary. These counts, are carried out when most of the preferred intertidal feeding habitats are temporarily unavailable to the species, with most either foraging on the upper shore in sub-optimal areas or roosting in the area while waiting for the tide to retreat. A certain site fidelity of the species was assumed, hence considering the obtained density data as representative of the bird use of the area, as most species tend to simply move up and/or along shore during tidal compression (and depending on tide), with roosts usually located as close to preferred foraging areas as possible. However, it must be acknowledged that the use of high-tide counts might lead to an underestimation of the birds using an area at low tide, particularly for those species (e.g. Redshank, Curlew and Oystercatcher) which can move inland in search of food when their estuarine food resources are not accessible (Prater 1981). Using low-tide counts is likely to allow better relationships of the actual density with the habitat availability and characteristics in the counting unit areas, although this type of data might present other limitations compared to the high-tide counts. For example, in the Humber, low-tide counts are carried out at a lower frequency (e.g. c. every 5 years) hence limiting the availability of data for the statistical analysis.
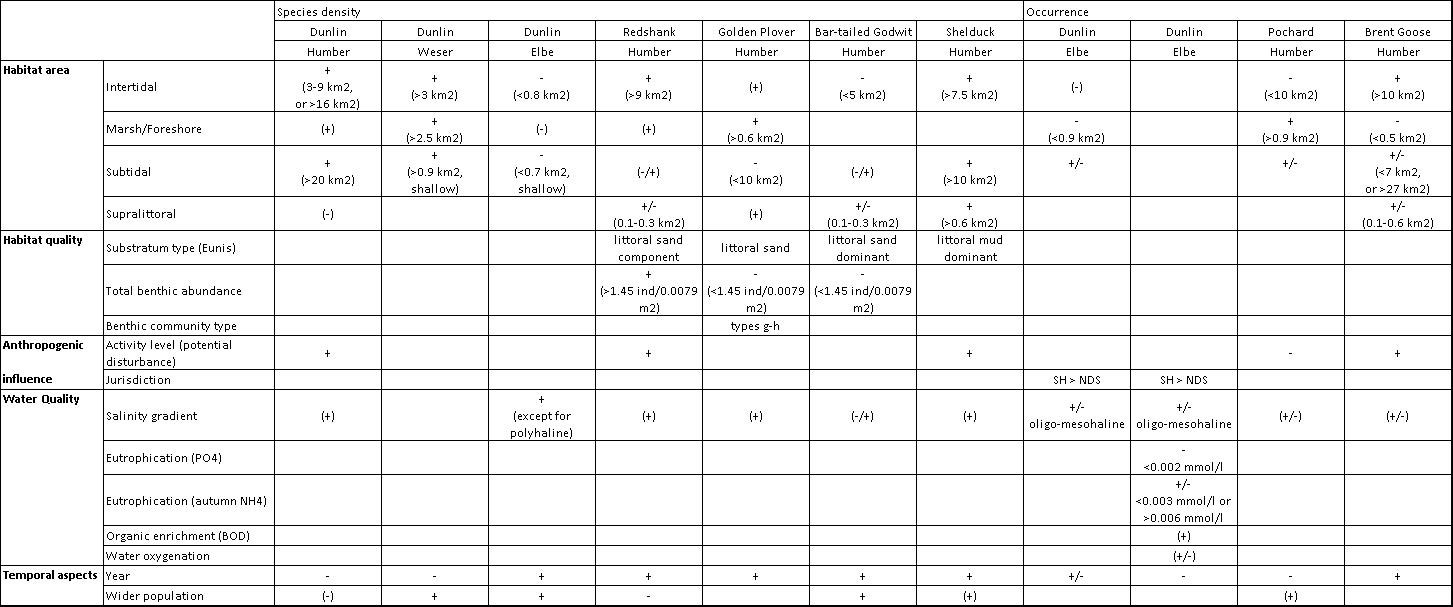
Symbols + and – indicate a general positive or negative relationship with the relevant environmental descriptor (thresholds and ranges identifying optimal conditions for the species occurrence/density are provided in brackets). +/- indicates a fluctuating relationship (when no clear increase or decrease pattern can be identified; the shape of these relationships being shown in Chapter 6). When symbols are within brackets, these indicate predictors that have not been included in the models because of their correlation with other variables included, but, due to their collinearity with the relevant descriptors, their influence cannot be excluded.
Important to know
Reports / Measures / Tools
| Report: | Management measures analysis and comparison |
|---|
Management issues
How can management targets and monitoring strategies be set for waterbirds in compensatory areas?What environmental factors should be considered in the design of a compensation scheme for waterbirds and their habitats?
What environmental variables are most important in determining minimizing or basic compensatory requirements for waterbirds?
What is important in establishing a zonation for estuaries?
What tools and guidance are available to minimise and mitigate disturbance to waterbirds?